Research history
Synthetic Biology is a field that employs engineering principles to construct new genetic systems. The approach is based on the use of well characterised and reusable DNA components, and numerical models for the design of biological circuits. The approach shows great potential for the engineering of multicellular systems, and plants are the obvious first target for this type of approach. Plants possess indeterminate and modular body plans, have a wide spectrum of biosynthetic activities, can be genetically manipulated, and are used globally for low cost bioproduction at up to gigatonne scale. However, modern approaches to engineering only became feasible after the development of foundational molecular genetic techniques and systems. I started my research career in the late 1970's - at a time when Molecular Biology was displacing more traditional Biochemistry approaches, and have been in a position to see another paradigm shift as Synthetic Biology and Engineering inspired ways of working started to take over in the early 2000's. The research work of my colleagues and I is outlined below, carried on these waves of innovation and creation of new tools.
Over time, it has become possible to tackle challenges at ever larger scale. Moving from molecular-scale analysis - to engineering molecules, to visualising living processes inside intact organisms and beginning to integrate and reprogram the physico-genetic interactions that underpin emergent growth processes. We are now seeing the arrival of the plant Marchantia polymorpha as a simple testbed for engineering cellular growth and metabolism. Marchantia is am early divergent descendant of the earliest terrestrial plants, and characterised by morphological simplicity, a highly streamlined genome, and a growing set of techniques for facile genetic manipulation, culture and microscopy. Marchantia is providing a new platform for reprogramming of plant development and physiology.
Over time, it has become possible to tackle challenges at ever larger scale. Moving from molecular-scale analysis - to engineering molecules, to visualising living processes inside intact organisms and beginning to integrate and reprogram the physico-genetic interactions that underpin emergent growth processes. We are now seeing the arrival of the plant Marchantia polymorpha as a simple testbed for engineering cellular growth and metabolism. Marchantia is am early divergent descendant of the earliest terrestrial plants, and characterised by morphological simplicity, a highly streamlined genome, and a growing set of techniques for facile genetic manipulation, culture and microscopy. Marchantia is providing a new platform for reprogramming of plant development and physiology.
Viroid RNA sequencing
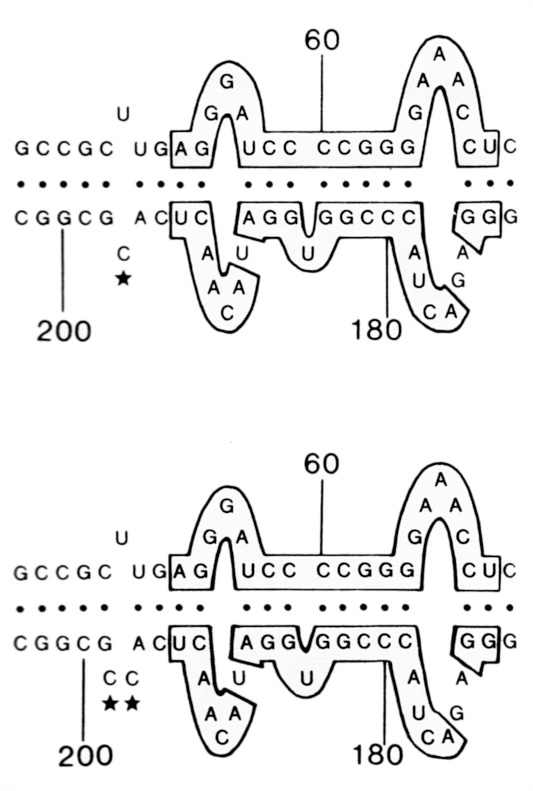
As a graduate student in the Laboratory of Prof. Robert Symons, University of Adelaide, Australia, I developed a new method for direct sequencing of circular RNAs, and used this to determine the complete genome of a number of viroid species. This work allowed us to identify the first conserved sequences within viroid genomes. This led to the characterisation of autocatalytic domains within the RNAs.
Haseloff, J. and Symons, R.H. Nuc. Acids Res. 9, 2741 (1981)
Haseloff, J. and Symons, R.H. Nuc. Acids Res. 10, 3681 (1982)
Haseloff, J., Mohamed, N.A. and Symons, R.H. Nature 299, 316 (1982).
Haseloff, J. and Symons, R.H. Nuc. Acids Res. 9, 2741 (1981)
Haseloff, J. and Symons, R.H. Nuc. Acids Res. 10, 3681 (1982)
Haseloff, J., Mohamed, N.A. and Symons, R.H. Nature 299, 316 (1982).
Plant virus expression
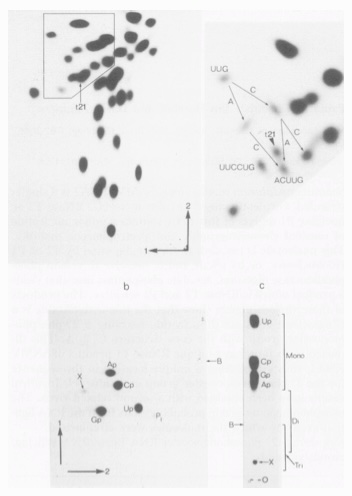
In 1983, I moved to work with Dr. Dave Zimmern at the MRC Laboratory of Molecular Biology, Cambridge. The first few complete sequences of RNA plant viruses had just been determined. Using early tools for bioinformatic analysis, we found the first similarities between these animal and plant viruses, that previously were thought to be unrelated. It is now recognised that these RNA viruses form a superfamily, probably due to horizontal transfer of genes between kingdoms. In addition, we discovered a chemical signature for the ligation of circular viroid-like molecules. An unusual 2’ phosphomonoester group is left as a relic of ligation, and indicates that the RNAs exploit a host tRNA ligase during replication.
Haseloff, J., Goelet, P., Zimmern, D., Alquist, P., Dasgupta, R. and Kaesberg, P. Proc. Natl. Acad. Sci. USA. 81, 4358, (1984)
Kiberstis, P.A., Haseloff, J. and Zimmern, D. EMBO J. 4, 817 (1985).
Haseloff, J., Goelet, P., Zimmern, D., Alquist, P., Dasgupta, R. and Kaesberg, P. Proc. Natl. Acad. Sci. USA. 81, 4358, (1984)
Kiberstis, P.A., Haseloff, J. and Zimmern, D. EMBO J. 4, 817 (1985).
Synthetic ribozymes
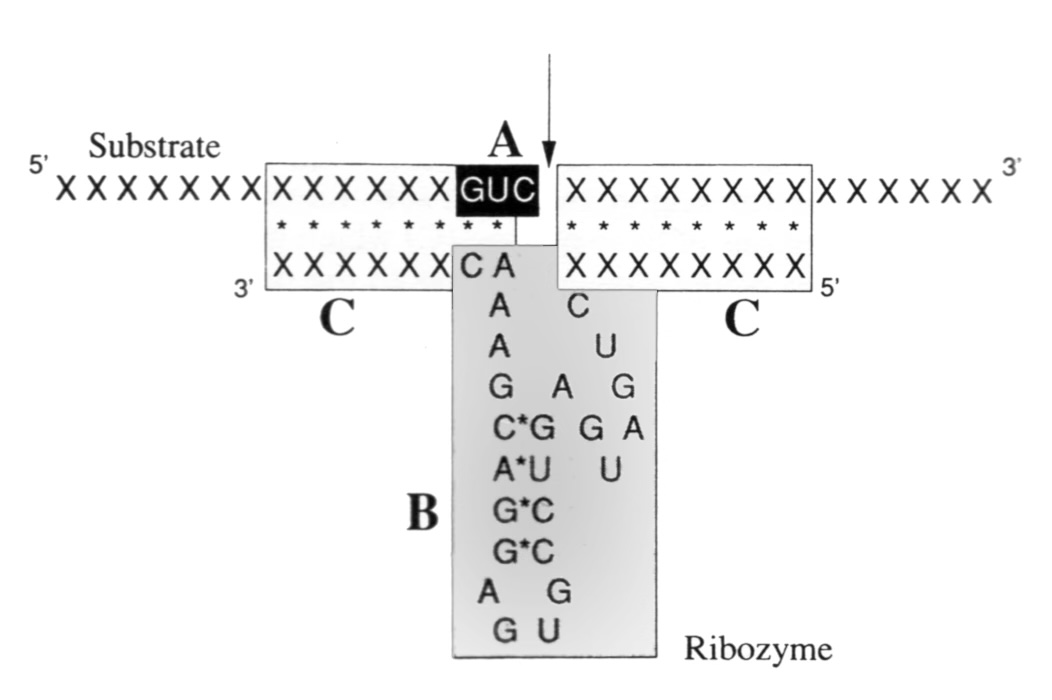
In 1985, I moved to the CSIRO Division of Plant Industry, Canberra, Australia. Working with Dr. Wayne Gerlach, I defined sequences required for the self-catalysed cleavage of a satellite virus RNA, and used this to describe the first simple design for artificial ribozymes with new substrate specificities. This was the first scheme for producing novel RNA enzymes at will. The published work triggered a great deal of scientific and commercial activity in attempts to use synthetic ribozymes as gene control agents in vivo. In the laboratory, we also experimented with new forms of virus resistance in transgenic plants, constructing a synthetic resistance gene from a satellite RNA replicon.
Gerlach, W.L., Llewellen, D. and Haseloff, J. Nature 328, 802 (1987)
Haseloff, J. and Gerlach, W.L. Nature 334, 585 (1988)
Haseloff, J. and Gerlach, W.L. Gene 82, 45 (1989).
Gerlach, W.L., Llewellen, D. and Haseloff, J. Nature 328, 802 (1987)
Haseloff, J. and Gerlach, W.L. Nature 334, 585 (1988)
Haseloff, J. and Gerlach, W.L. Gene 82, 45 (1989).
Arabidopsis engineering

In 1989, I moved to the Department of Molecular Biology, Harvard Medical School, to work with Prof. Howard Goodman. I was very keen to learn molecular genetic techniques for working with Arabidopsis, which was then an emerging system for studying plant development. I began development of the GAL4 system in plants, and described the development and testing of novel trans-splicing ribozymes that can be used as RNA sensors and delivery agents.
Kohler, U. Ayre, B., Goodman, H. M. and Haseloff, J. J. Mol. Biol. 284, 1935-1950 (1999)
Ayre, B.G., Köhler, U., Goodman, H.M. and Haseloff, J. Proc. Natl. Acad. Sci. USA. 96, 3507-3512 (1999).
Kohler, U. Ayre, B., Goodman, H. M. and Haseloff, J. J. Mol. Biol. 284, 1935-1950 (1999)
Ayre, B.G., Köhler, U., Goodman, H.M. and Haseloff, J. Proc. Natl. Acad. Sci. USA. 96, 3507-3512 (1999).
Visualising plant gene expression

This work continued after 1993, when I returned to Cambridge, to establish a group at the MRC Laboratory of Molecular Biology. Our small laboratory turned to the development of tools for engineering plant development, including a battery of novel microscopic, genetic and computational tools for visualising and manipulating gene expression in live plants.
We were the first to use obtain and use the green fluorescent protein in plants. We identified and cured a cryptic intron in the natural gene for green fluorescent protein (GFP), and produced modified forms of GFP with improved properties. These genes found wide use among plant scientists. We completed the construction and characterisation of a library of Arabidopsis plant lines expressing different patterns of a foreign transcription activator, GAL4, and GFP that provide high resolution cellular markers and can be used for precise misexpression studies. These were widely distributed through the Arabidopsis stock centres.
Siemering, K. R., Golbik, R., Sever, R. and Haseloff, J. Current Biology 6, 1653-1663, (1996)
Haseloff, J., Siemering, K. R., Prasher, D.C. and Hodge, S. Proc. Natl. Acad. Sci. USA. 94, 2122-2127 (1997).
Haseloff, J, Methods in Cell Biology, Vol. 58 (1999).
We were the first to use obtain and use the green fluorescent protein in plants. We identified and cured a cryptic intron in the natural gene for green fluorescent protein (GFP), and produced modified forms of GFP with improved properties. These genes found wide use among plant scientists. We completed the construction and characterisation of a library of Arabidopsis plant lines expressing different patterns of a foreign transcription activator, GAL4, and GFP that provide high resolution cellular markers and can be used for precise misexpression studies. These were widely distributed through the Arabidopsis stock centres.
Siemering, K. R., Golbik, R., Sever, R. and Haseloff, J. Current Biology 6, 1653-1663, (1996)
Haseloff, J., Siemering, K. R., Prasher, D.C. and Hodge, S. Proc. Natl. Acad. Sci. USA. 94, 2122-2127 (1997).
Haseloff, J, Methods in Cell Biology, Vol. 58 (1999).
High resolution imaging
In March 1999, I took up a Sainsbury Fellowship, and my laboratory moved to the Department of Plant Sciences, University of Cambridge. I obtained funding to establish new confocal microscopy and computing facilities, and we have developed improved microscopy and software methods for the analysis and engineering of cell dynamics during morphogenesis.
We used the GAL4-GFP Arabidopsis lines as high resolution cellular markers for both developmental and physiological studies. Given the great utility of the lines, we produced a second library of lines using a complementary enhancer trap vector based on HAP1, a close relative of GAL4, and a cyan fluorescent protein - histone 2b marker gene. We have developed ratiometric techniques for cell-scale quantification of gene expression in live plants and bacteria. This allows the extraction of parameters for computational modelling of biological systems at cellular resolution.
Bougourd, S., Marrison, J. and Haseloff, J. Plant Journal, 24, 543-550 (2000)
Haseloff J. Biotechniques. 34:1174-1182 (2003)
Berger, F., Linstead, P., Dolan, L. and Haseloff, J. Developmental Biology 194, 226-234, (1998)
Berger, F., Haseloff, J., Schiefelbein, J. and Dolan, L.. Current Biology 8, 421-430 (1998)
Boisnard-Lorig, C., et al., Plant Cell 13: 495-509 (2001).
Kurup, S., Runions, J.C., Kohler, U., Laplaze, L., Hodge, S. and Haseloff, J. Plant Journal, 42:444-453 (2005).
Coates, J.C., Laplaze, L. and Haseloff, J. Proc. Natl. Acad Sci. USA, 103:1621-1626 (2006).
Truernit E, Haseloff J. Plant Methods. 23;4:15 (2008).
Federici F, Dupuy L, Laplaze L, Heisler M & Haseloff J. Nature Methods, 9:483-485 (2012)
We used the GAL4-GFP Arabidopsis lines as high resolution cellular markers for both developmental and physiological studies. Given the great utility of the lines, we produced a second library of lines using a complementary enhancer trap vector based on HAP1, a close relative of GAL4, and a cyan fluorescent protein - histone 2b marker gene. We have developed ratiometric techniques for cell-scale quantification of gene expression in live plants and bacteria. This allows the extraction of parameters for computational modelling of biological systems at cellular resolution.
Bougourd, S., Marrison, J. and Haseloff, J. Plant Journal, 24, 543-550 (2000)
Haseloff J. Biotechniques. 34:1174-1182 (2003)
Berger, F., Linstead, P., Dolan, L. and Haseloff, J. Developmental Biology 194, 226-234, (1998)
Berger, F., Haseloff, J., Schiefelbein, J. and Dolan, L.. Current Biology 8, 421-430 (1998)
Boisnard-Lorig, C., et al., Plant Cell 13: 495-509 (2001).
Kurup, S., Runions, J.C., Kohler, U., Laplaze, L., Hodge, S. and Haseloff, J. Plant Journal, 42:444-453 (2005).
Coates, J.C., Laplaze, L. and Haseloff, J. Proc. Natl. Acad Sci. USA, 103:1621-1626 (2006).
Truernit E, Haseloff J. Plant Methods. 23;4:15 (2008).
Federici F, Dupuy L, Laplaze L, Heisler M & Haseloff J. Nature Methods, 9:483-485 (2012)
Cell modelling

For any attempt to engineer plant cell interactions, it is necessary to have a computational framework for modelling system dynamics. We generated and tested a software description of the physical and genetic properties of cells. CellModeller contained a physico-genetic model that was GPU accelerated and provided an engine for the in silico growth of large populations of plant or microbial cells through enlargement and division. Fields of proliferating cells could be programmed via a genetic script to produce and respond to different genetic programmes. We used CellModeller to explore prototypes of simple artificial regulatory networks that could be implemented to implement synthetic patterning systems in vivo.
Rudge, T. and Haseloff, J. Lecture Notes in Computer Science: Advances in Artificial Life, 3630:78-87 (2005).
Dupuy, L.X., Mackenzie, J and Haseloff, J. Proceedings of the 5th Plant Biomechanics Conference. Stockholm. (2006)
Dupuy L, Mackenzie J, Rudge T, Haseloff J. Annals of Botany 101:1255-1265 (2008).
Dupuy, L., Mackenzie, J. and Haseloff, J. Proc. Natl. Acad. Sci. USA 107:2711-6 (2010).
Rudge TJ, Steiner PJ, Phillips A and Haseloff J. ACS Synthetic Biology, 1:345-352, (2012).
Rudge, T. and Haseloff, J. Lecture Notes in Computer Science: Advances in Artificial Life, 3630:78-87 (2005).
Dupuy, L.X., Mackenzie, J and Haseloff, J. Proceedings of the 5th Plant Biomechanics Conference. Stockholm. (2006)
Dupuy L, Mackenzie J, Rudge T, Haseloff J. Annals of Botany 101:1255-1265 (2008).
Dupuy, L., Mackenzie, J. and Haseloff, J. Proc. Natl. Acad. Sci. USA 107:2711-6 (2010).
Rudge TJ, Steiner PJ, Phillips A and Haseloff J. ACS Synthetic Biology, 1:345-352, (2012).
Experimental microbial models
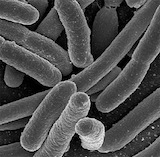
We used microbial biofilms as surrogate plant cell populations in order to construct synthetic patterning systems for breaking symmetry The combination of computational models, quantitative microscopy and simple experimental systems for cell dynamics allowed direct manipulation and insight into the interplay between genetic and physical systems. Even simple cellular programming of growth, cohort recruitment and boundary reinforcement lead to emergent behaviour.
Rudge TJ, Steiner PJ, Kan A and Haseloff J. ACS Synthetic Biology, DOI: 10.1021/sb400030p (2013).
Rudge T, Brown J, Federici F, Dalchau N, Phillips A, Ajioka J, & Haseloff J. ACS Synthetic Biology Jan 5:89-98. (2016).
Grant PK, Dalchau N, Brown JR, Federici F, Rudge TJ, Yordanov B, Patange O, Phillips A, Haseloff J. Molecular Systems Biology 12:849-861, (2016).
Anton Kan, Ilenne Del Valle, Tim Rudge, Fernan Federici and Jim Haseloff. Journal of The Royal Society Interface. doi.org/10.1098/rsif.2018.0406, (2018).
CR Boehm, PK Grant and J Haseloff. Nature Communications, 9: 776 (2018).
Rudge TJ, Steiner PJ, Kan A and Haseloff J. ACS Synthetic Biology, DOI: 10.1021/sb400030p (2013).
Rudge T, Brown J, Federici F, Dalchau N, Phillips A, Ajioka J, & Haseloff J. ACS Synthetic Biology Jan 5:89-98. (2016).
Grant PK, Dalchau N, Brown JR, Federici F, Rudge TJ, Yordanov B, Patange O, Phillips A, Haseloff J. Molecular Systems Biology 12:849-861, (2016).
Anton Kan, Ilenne Del Valle, Tim Rudge, Fernan Federici and Jim Haseloff. Journal of The Royal Society Interface. doi.org/10.1098/rsif.2018.0406, (2018).
CR Boehm, PK Grant and J Haseloff. Nature Communications, 9: 776 (2018).
The Marchantia system

These approaches have led to the adoption of the liverwort plant Marchantia polymorpha as a new vehicle for genome-scale manipulation. Unlike seed plants, it forms spores, and can be handled as single cells, where targeted gene modifications, plastid transformation, regeneration, high throughput optical analysis techniques are highly efficient. It bridges the gap between microbial and plant systems.
John L. Bowman et al, Cell, 171:287–304 (2017), including:
Bernardo Pollak, Mihails Delmans and Jim Haseloff, Supplementary material.
Romani F, Sauret-Güeto S, Rebmann M, Annese D, Bonter I, Tomaselli M, Dierschke T, Delmans M, Frangedakis E, Silvestri L, Rever J, Bowman JL, Romani I, Haseloff J. Plant Cell. 36(6):2140-2159. doi: 10.1093/plcell/koae053. PMID: 38391349 (2024)
Forestier ECF, Asprilla P, Bonter I, Romani F, Frangedakis E, Haseloff J. Communications Biology 9(1):521. doi: 10.1038/s42003-025-09508-4. (2026)
Romani F, Bonter I, Rebmann M, Takahashi G, Guzman-Chavez F, De Batté F, Hirakawa Y, Haseloff J. Plant Cell 38(6):koag103. doi: 10.1093/plcell/koag103. (2026)
John L. Bowman et al, Cell, 171:287–304 (2017), including:
Bernardo Pollak, Mihails Delmans and Jim Haseloff, Supplementary material.
Romani F, Sauret-Güeto S, Rebmann M, Annese D, Bonter I, Tomaselli M, Dierschke T, Delmans M, Frangedakis E, Silvestri L, Rever J, Bowman JL, Romani I, Haseloff J. Plant Cell. 36(6):2140-2159. doi: 10.1093/plcell/koae053. PMID: 38391349 (2024)
Forestier ECF, Asprilla P, Bonter I, Romani F, Frangedakis E, Haseloff J. Communications Biology 9(1):521. doi: 10.1038/s42003-025-09508-4. (2026)
Romani F, Bonter I, Rebmann M, Takahashi G, Guzman-Chavez F, De Batté F, Hirakawa Y, Haseloff J. Plant Cell 38(6):koag103. doi: 10.1093/plcell/koag103. (2026)
Systematic tools for plant engineering
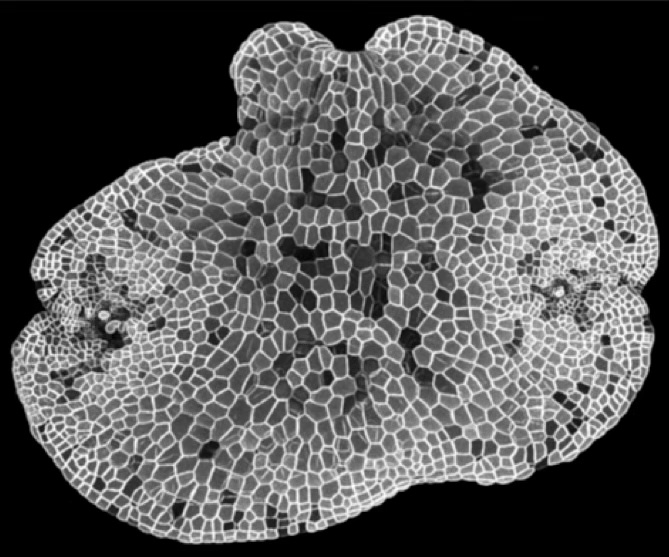
The molecular genetic tools that we have developed in Arabidopsis are being transferred to Marchantia. In addition we have established new community standards for DNA parts (Phytobricks), Loop assembly for efficient automated DNA construction and the OpenMTA as a legal framework for global sharing and creation of a commons for free exchange of DNA parts. Marchantia and these engineering frameworks are becoming increasingly important for foundational Synthetic Biology studies in plants. The open sharing of tools for plant synthetic biology was promoted under OpenPlant (https://www.openplant.org).
Patron NJ et al. New Phytologist 208:13-9. (2015).
Bowman JL, Araki T, Arteaga-Vazquez MA, Berger F, Dolan L, Haseloff J, Ishizaki K, Kyozuka J, Lin SS, Nagasaki H, Nakagami H, Nakajima K, Nakamura Y, Ohashi-Ito K, Sawa S, Shimamura M, Solano R, Tsukaya H, Ueda T, Watanabe Y, Yamato KT, Zachgo S, Kohchi T. Plant Cell Physiol. 57:257-61, (2016).
Delmans M, Pollak B and Haseloff J, Plant Cell Physiol. 58: e5(1–9) (2016).
Pollak B, Cerda A, Delmans M, Álamos S, Moyano T, West A, Gutiérrez RA, Patron N, Federici F, Haseloff J. New Phytologist, (2018).
Kahl L, Molloy J, Patron N, Matthewman C, Haseloff J, Grewal D, Johnson R, Endy D. Nature Biotechnology 36:923-927 (2018).
Sauret-Gueto S, Frangedakis E, Silvestri L, Rebmann M, Tomaselli M, Markel K, Delmans M, West A, Patron N, Haseloff J. ACS Synthetic Biology 9, 4, 864–882 (2020).
Guzman-Chavez F, Arce A, Adhikari A, Vadhin S, Pedroza-Garcia JA, Gandini C, Ajioka JW, Molloy J, Sanchez-Nieto S, Varner JD, Federici F, Haseloff J. ACS Synthetic Biology. 11(3):1114-1128, (2022).
Marron AO, Sauret-Güeto S, Rebmann M, Silvestri L, Tomaselli M, Haseloff J. Plant Journal 116(2):604-628, (2023).
Tse SW, Annese D, Romani F, Guzman-Chavez F, Bonter I, Forestier E, Frangedakis E, Haseloff J. Plant Cell Physiol. 65(8):1298-1309, (2024).
Annese D, Romani F, Grandellis C, Ives L, Frangedakis E, Buson FX, Molloy JC, Haseloff J. Plant Journal. 122(1):e70118. doi: 10.1111/tpj.70118. (2025)
Patron NJ et al. New Phytologist 208:13-9. (2015).
Bowman JL, Araki T, Arteaga-Vazquez MA, Berger F, Dolan L, Haseloff J, Ishizaki K, Kyozuka J, Lin SS, Nagasaki H, Nakagami H, Nakajima K, Nakamura Y, Ohashi-Ito K, Sawa S, Shimamura M, Solano R, Tsukaya H, Ueda T, Watanabe Y, Yamato KT, Zachgo S, Kohchi T. Plant Cell Physiol. 57:257-61, (2016).
Delmans M, Pollak B and Haseloff J, Plant Cell Physiol. 58: e5(1–9) (2016).
Pollak B, Cerda A, Delmans M, Álamos S, Moyano T, West A, Gutiérrez RA, Patron N, Federici F, Haseloff J. New Phytologist, (2018).
Kahl L, Molloy J, Patron N, Matthewman C, Haseloff J, Grewal D, Johnson R, Endy D. Nature Biotechnology 36:923-927 (2018).
Sauret-Gueto S, Frangedakis E, Silvestri L, Rebmann M, Tomaselli M, Markel K, Delmans M, West A, Patron N, Haseloff J. ACS Synthetic Biology 9, 4, 864–882 (2020).
Guzman-Chavez F, Arce A, Adhikari A, Vadhin S, Pedroza-Garcia JA, Gandini C, Ajioka JW, Molloy J, Sanchez-Nieto S, Varner JD, Federici F, Haseloff J. ACS Synthetic Biology. 11(3):1114-1128, (2022).
Marron AO, Sauret-Güeto S, Rebmann M, Silvestri L, Tomaselli M, Haseloff J. Plant Journal 116(2):604-628, (2023).
Tse SW, Annese D, Romani F, Guzman-Chavez F, Bonter I, Forestier E, Frangedakis E, Haseloff J. Plant Cell Physiol. 65(8):1298-1309, (2024).
Annese D, Romani F, Grandellis C, Ives L, Frangedakis E, Buson FX, Molloy JC, Haseloff J. Plant Journal. 122(1):e70118. doi: 10.1111/tpj.70118. (2025)
Chloroplast engineering

Chloroplasts are the main source of energy, reducing power and biosynthetic capacity in land plants. There may be 10-100 chloroplasts in a plant cell, and each plastid contains many copies (10-100) of the relatively small chloroplast genome (121 Kb in Marchantia). The plastid genome has high ploidy, is not subject to gene-silencing and capable of supporting hyperexpression of resident genes. Genetic transformation of chloroplasts is generally a protracted process and limited to few plant species. However the availability of spores and high regenerability in Marchantia makes this the most facile plant system for chloroplast transformation. We have developed and applied tools to establish this as a testbed for chloroplast engineering.
Boehm CR, Ueda M, Nishimura Y, Shikanai T, Haseloff J. Plant Cell Physiol. 57:291-9. (2016).
Frangedakis E, Markel K, Sauret-Gueto S, Haseloff J. Methods Mol Biol. 2317:343-365 (2021).
Frangedakis E, Guzman-Chavez F, Rebmann M, Markel K, Yu Y, Perraki A, Tse SW, Liu Y, Rever J, Sauret-Gueto S, Goffinet B, Schneider H, Haseloff J. ACS Synthetic Biology 16;10(7):1651-1666 (2021).
Frangedakis E, Yelina NE, Billakurthi K, Hua L, Schreier T, Dickinson PJ, Tomaselli M, Haseloff J, Hibberd JM. Cell. 187(18):4859-4876.e22. doi: 10.1016/j.cell.2024.06.039. Epub 2024 Jul 23. PMID: 39047726 (2024)
Boehm CR, Ueda M, Nishimura Y, Shikanai T, Haseloff J. Plant Cell Physiol. 57:291-9. (2016).
Frangedakis E, Markel K, Sauret-Gueto S, Haseloff J. Methods Mol Biol. 2317:343-365 (2021).
Frangedakis E, Guzman-Chavez F, Rebmann M, Markel K, Yu Y, Perraki A, Tse SW, Liu Y, Rever J, Sauret-Gueto S, Goffinet B, Schneider H, Haseloff J. ACS Synthetic Biology 16;10(7):1651-1666 (2021).
Frangedakis E, Yelina NE, Billakurthi K, Hua L, Schreier T, Dickinson PJ, Tomaselli M, Haseloff J, Hibberd JM. Cell. 187(18):4859-4876.e22. doi: 10.1016/j.cell.2024.06.039. Epub 2024 Jul 23. PMID: 39047726 (2024)